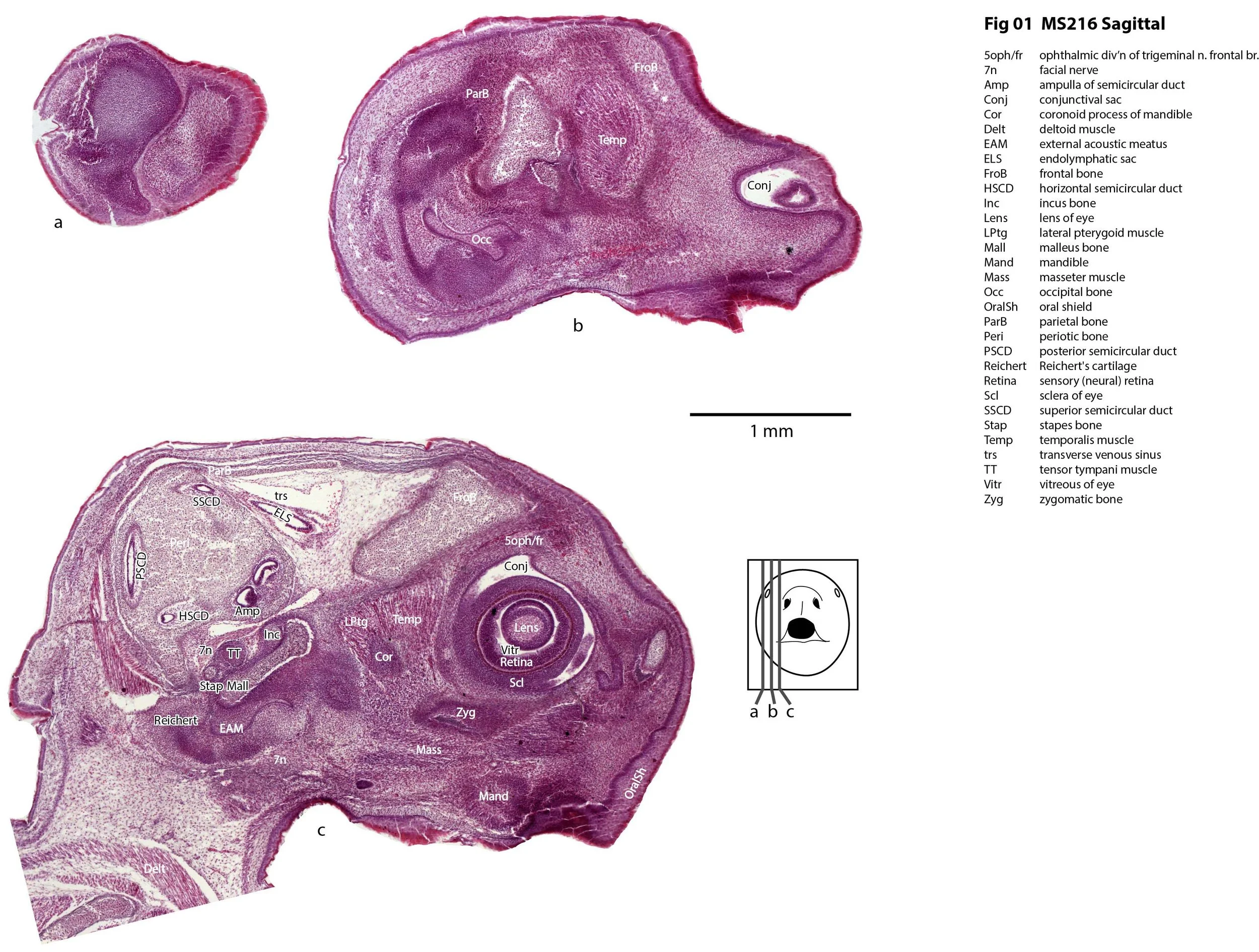
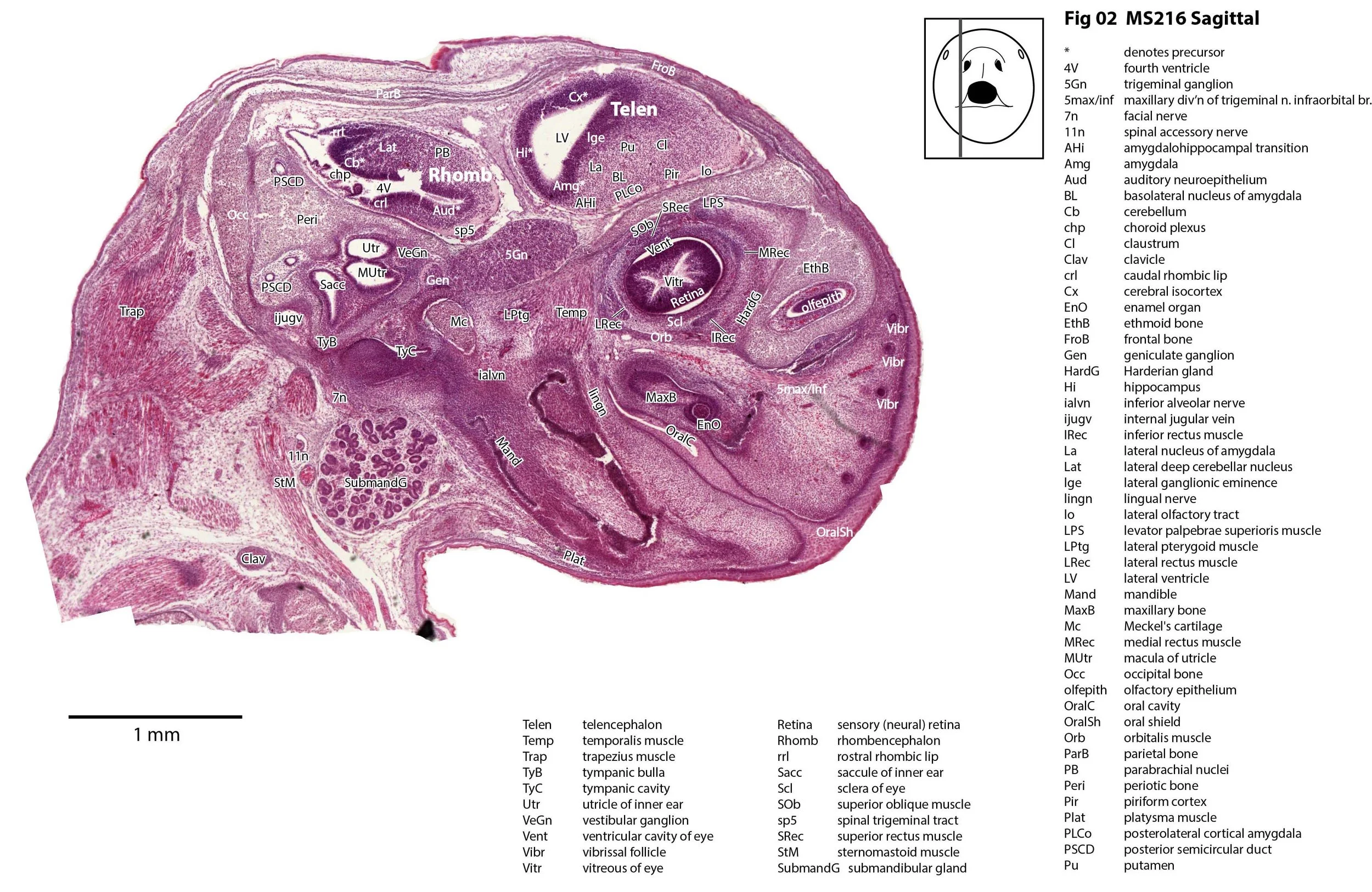
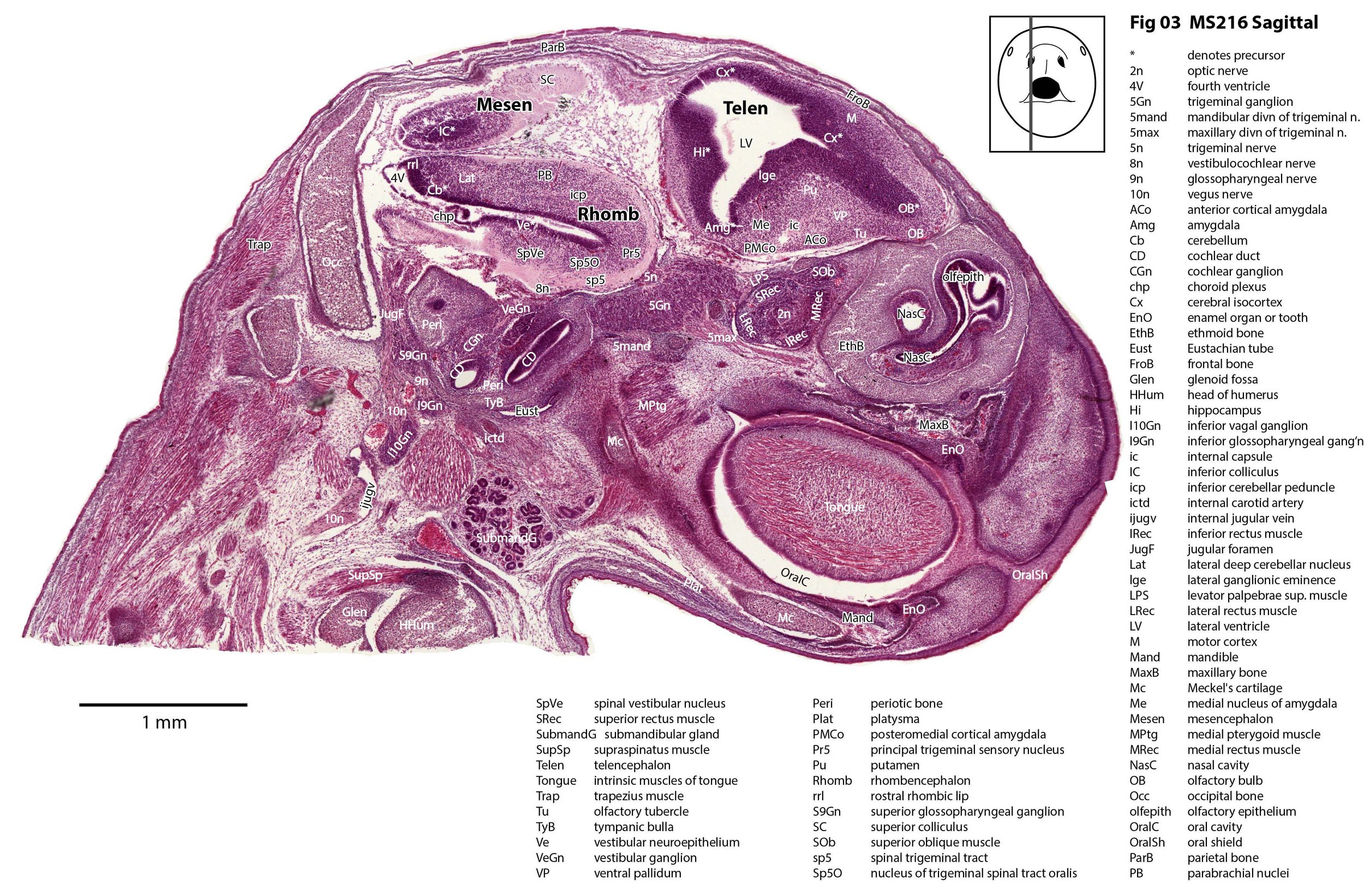
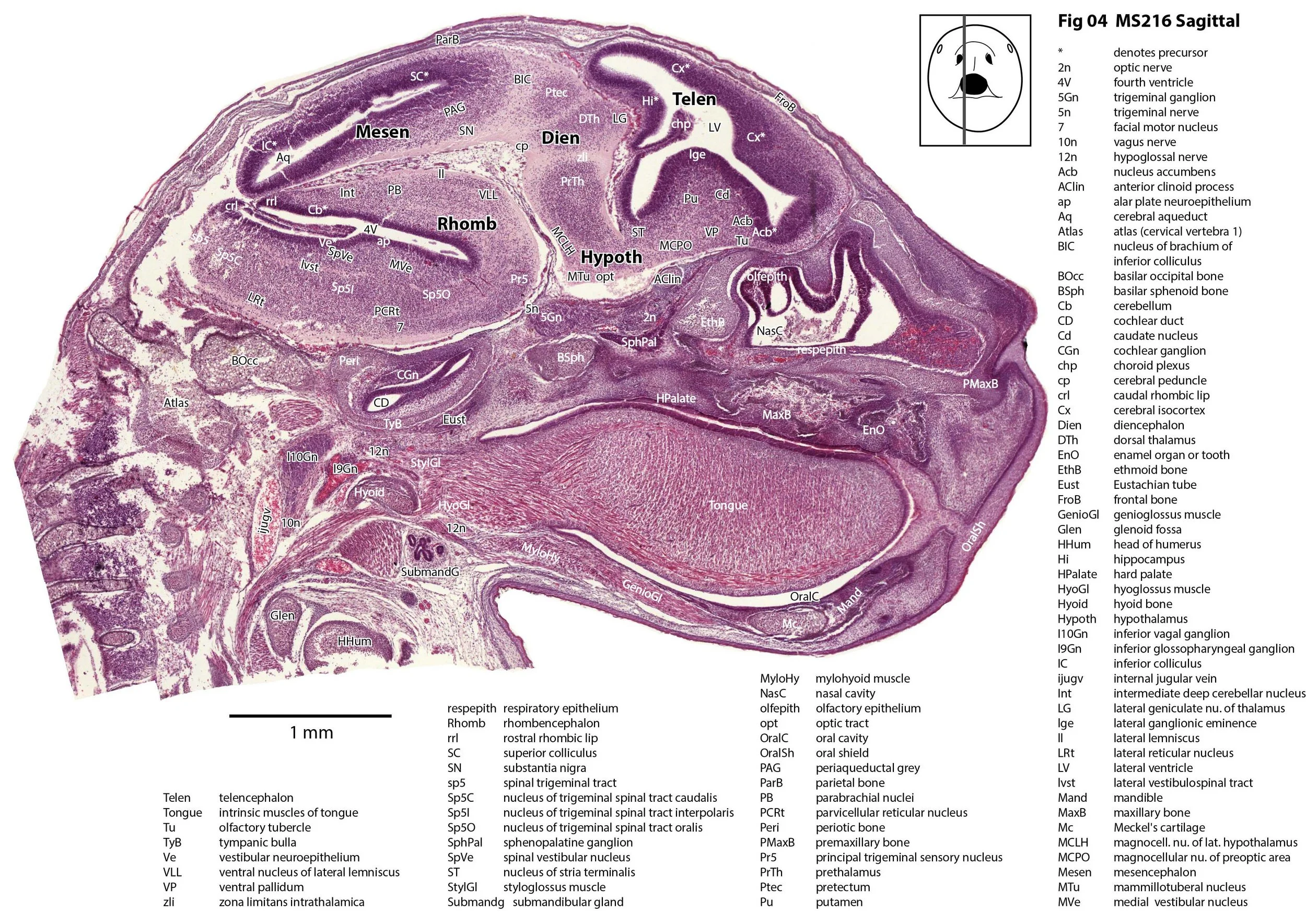
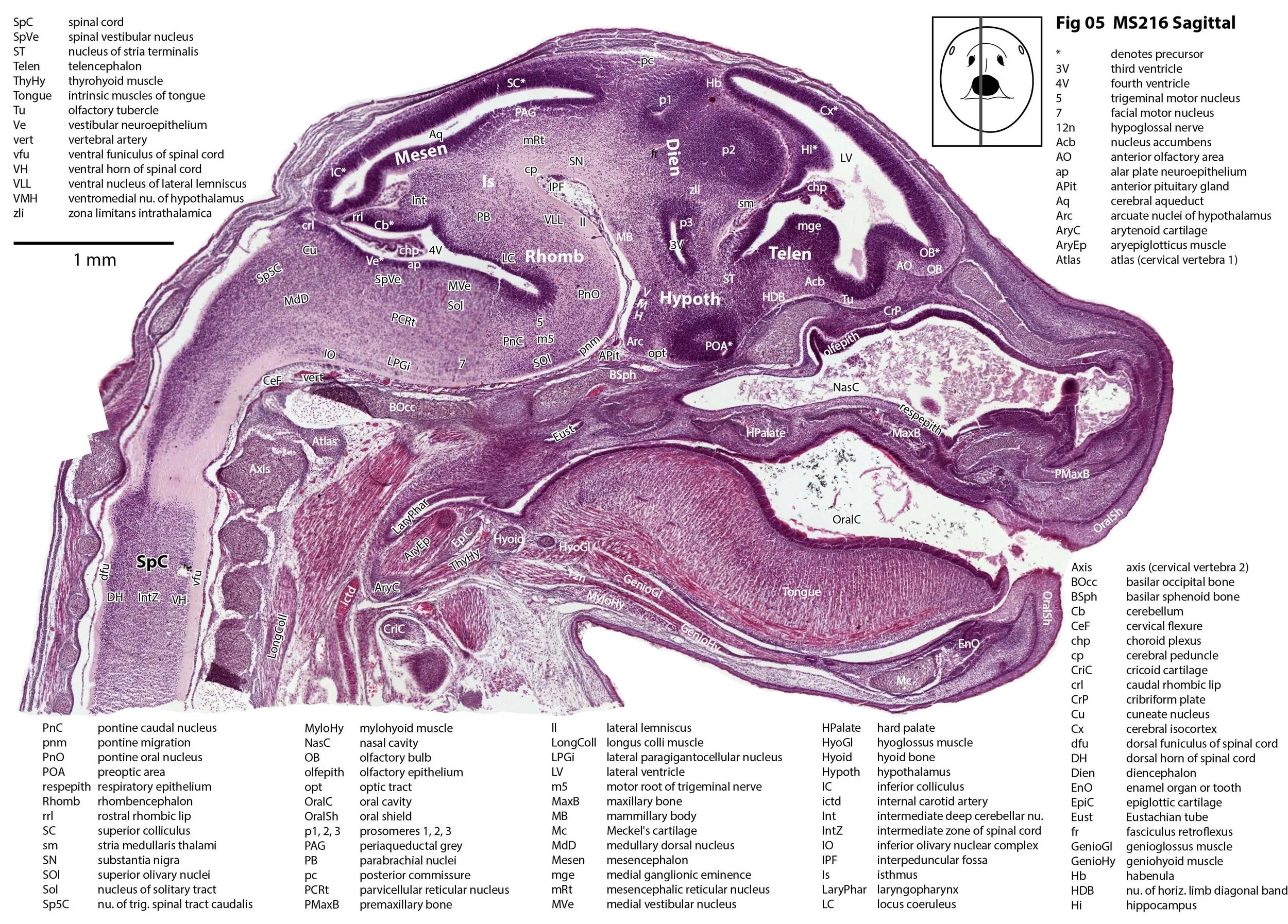
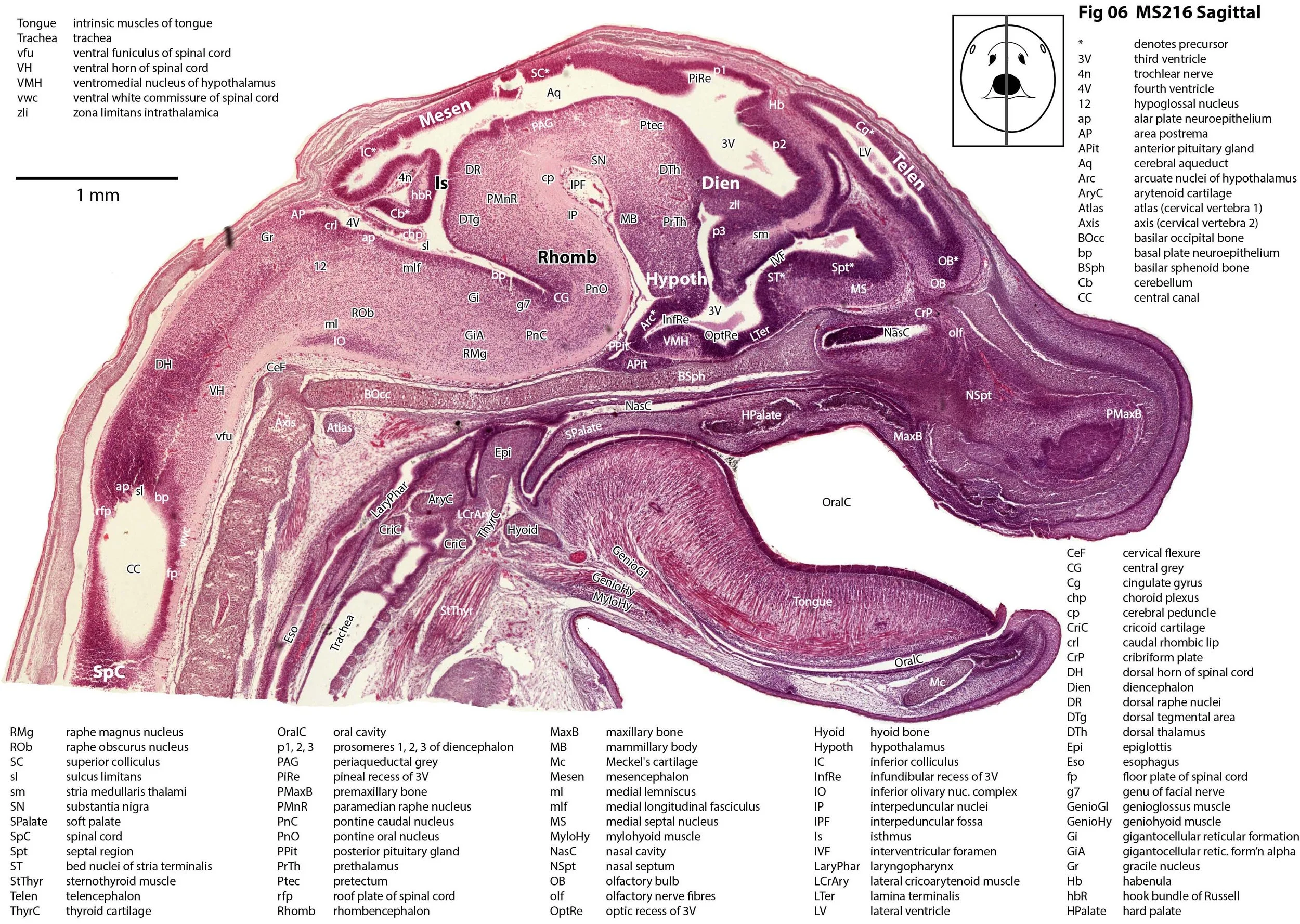
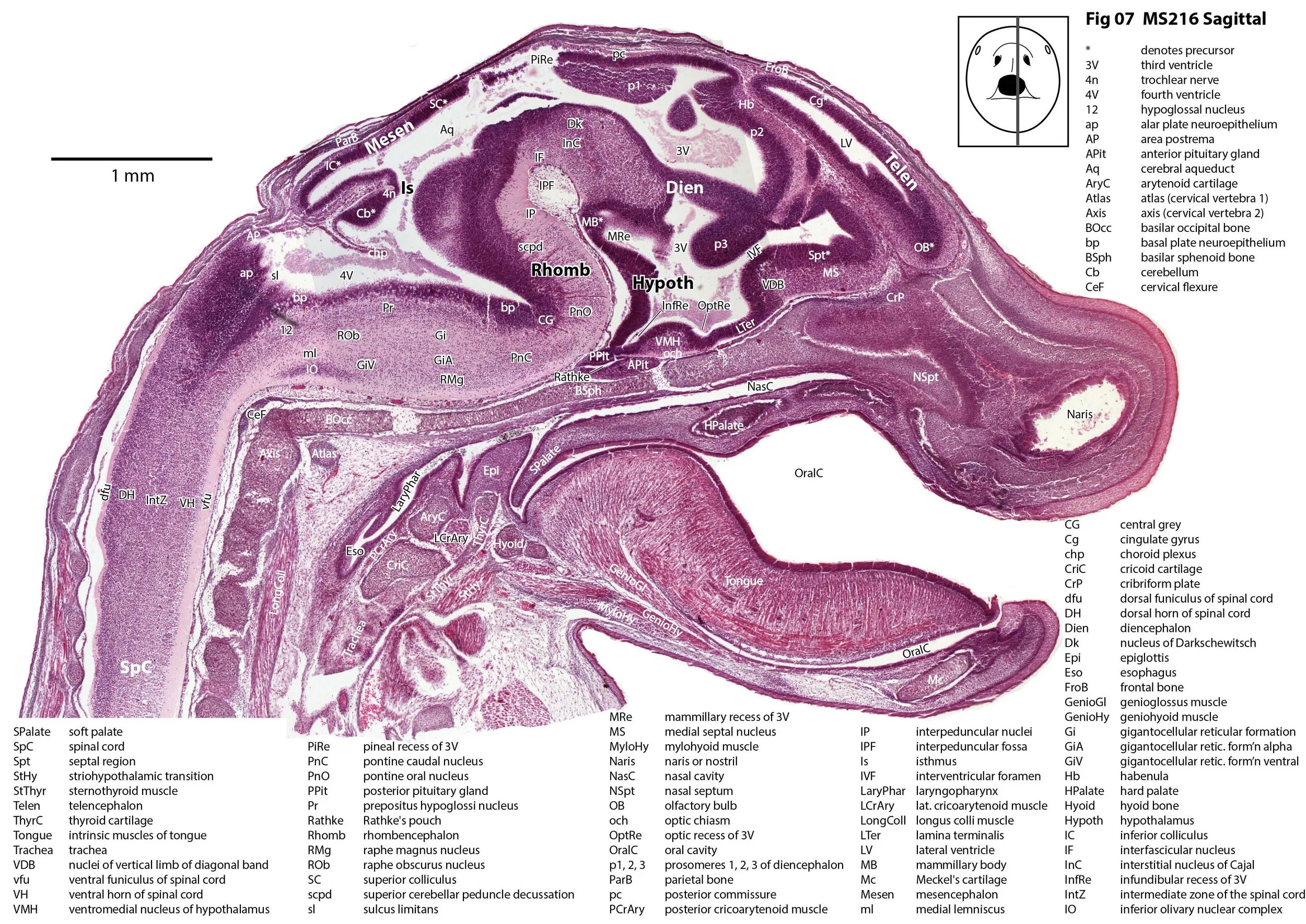
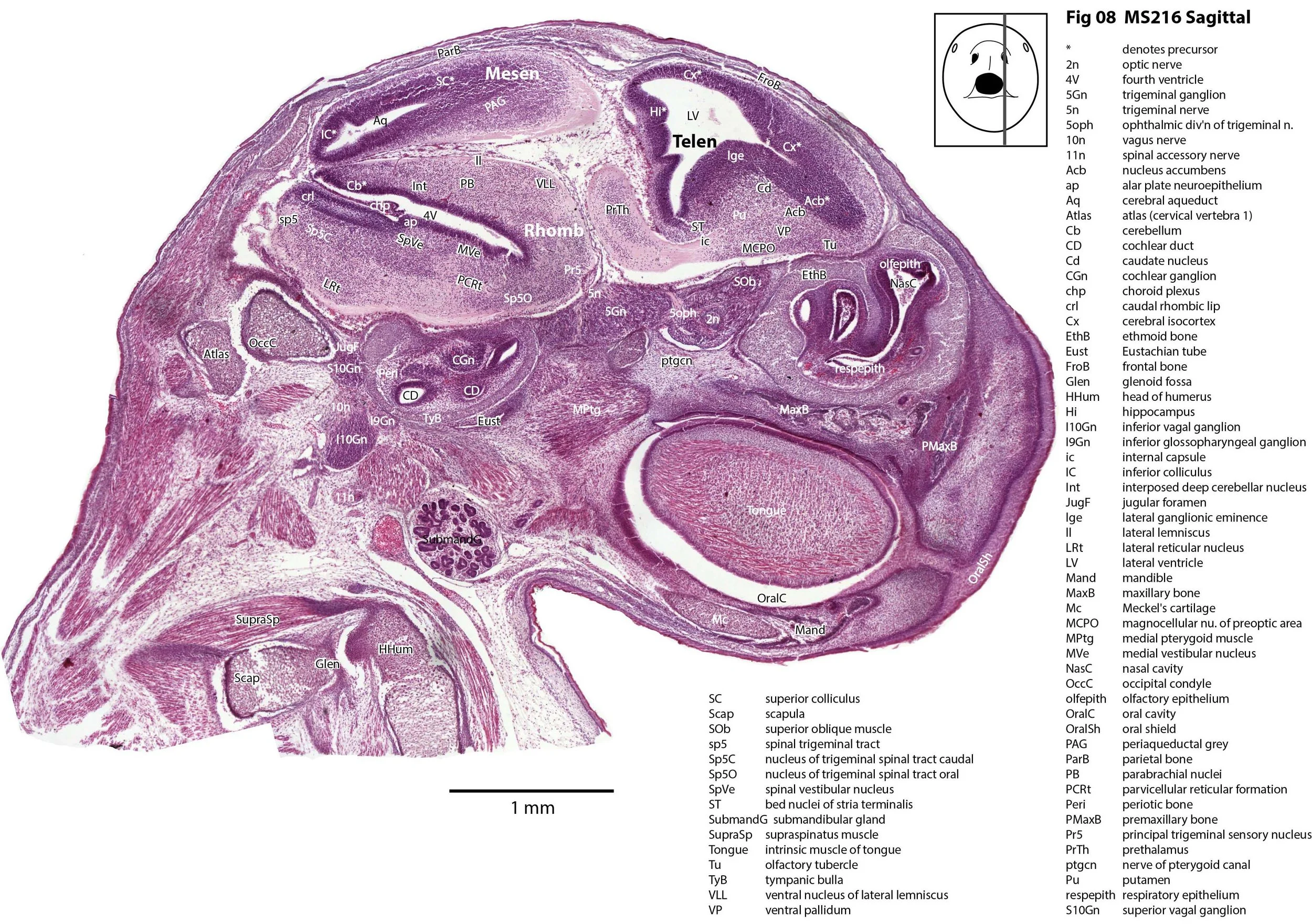
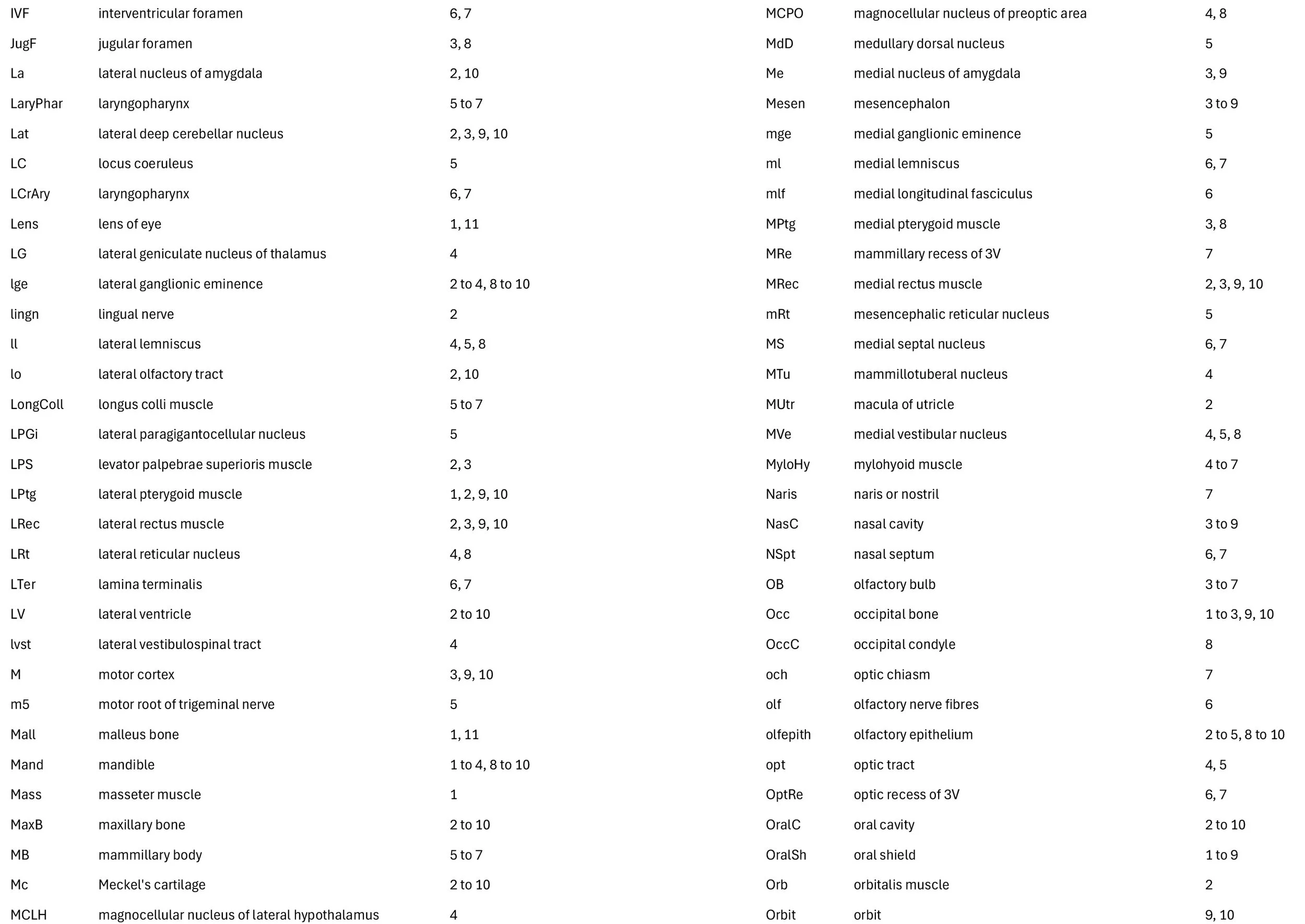
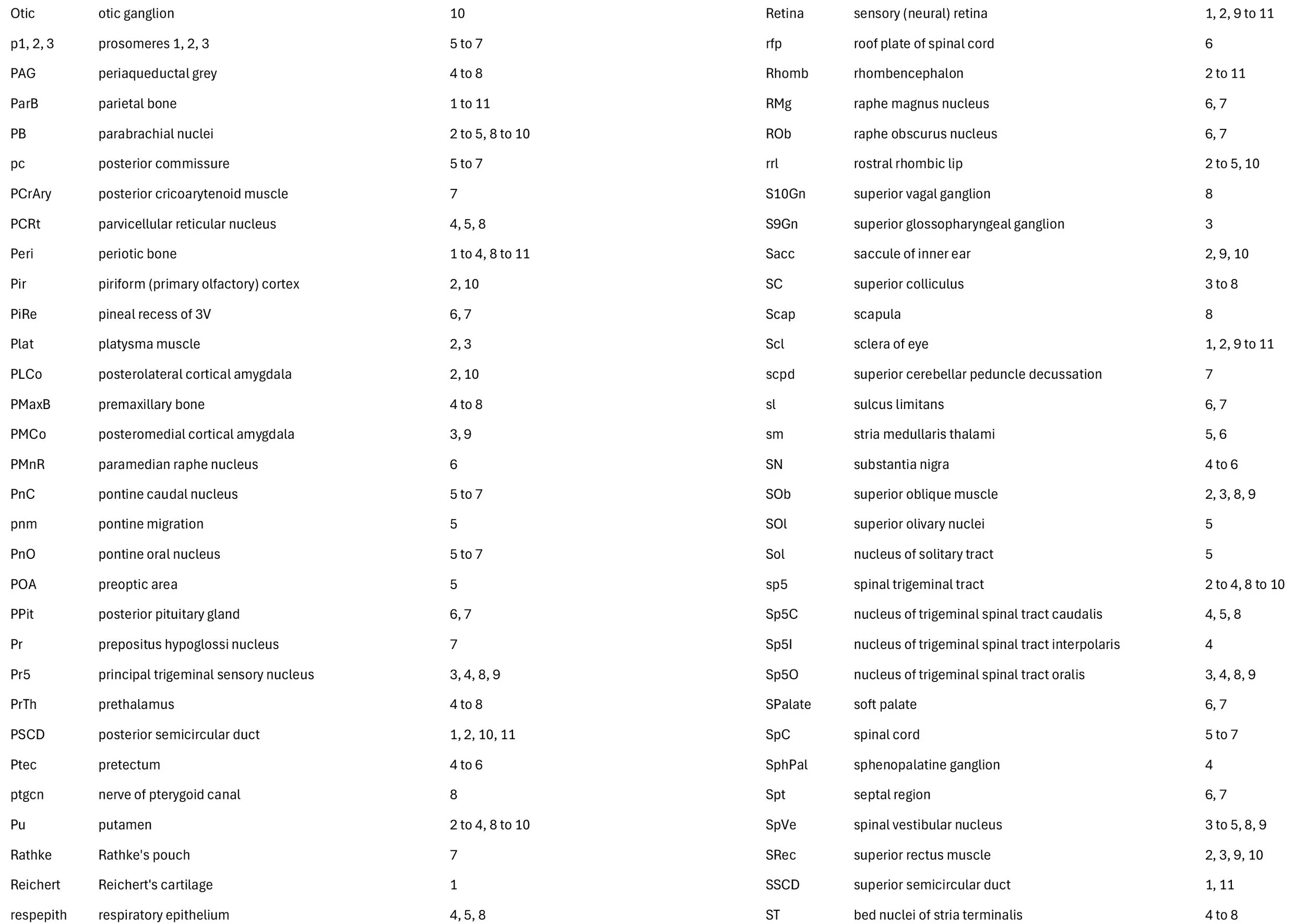
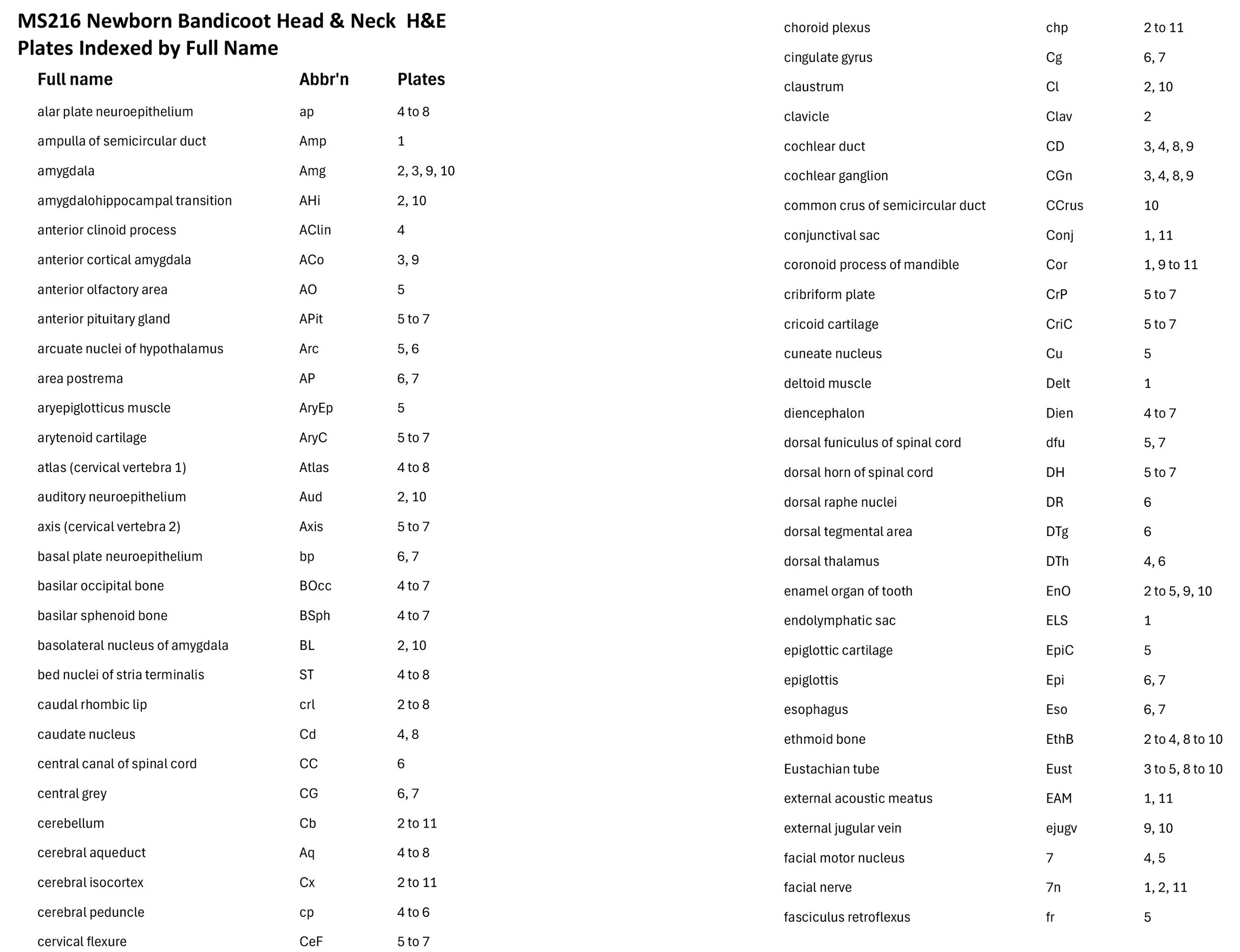
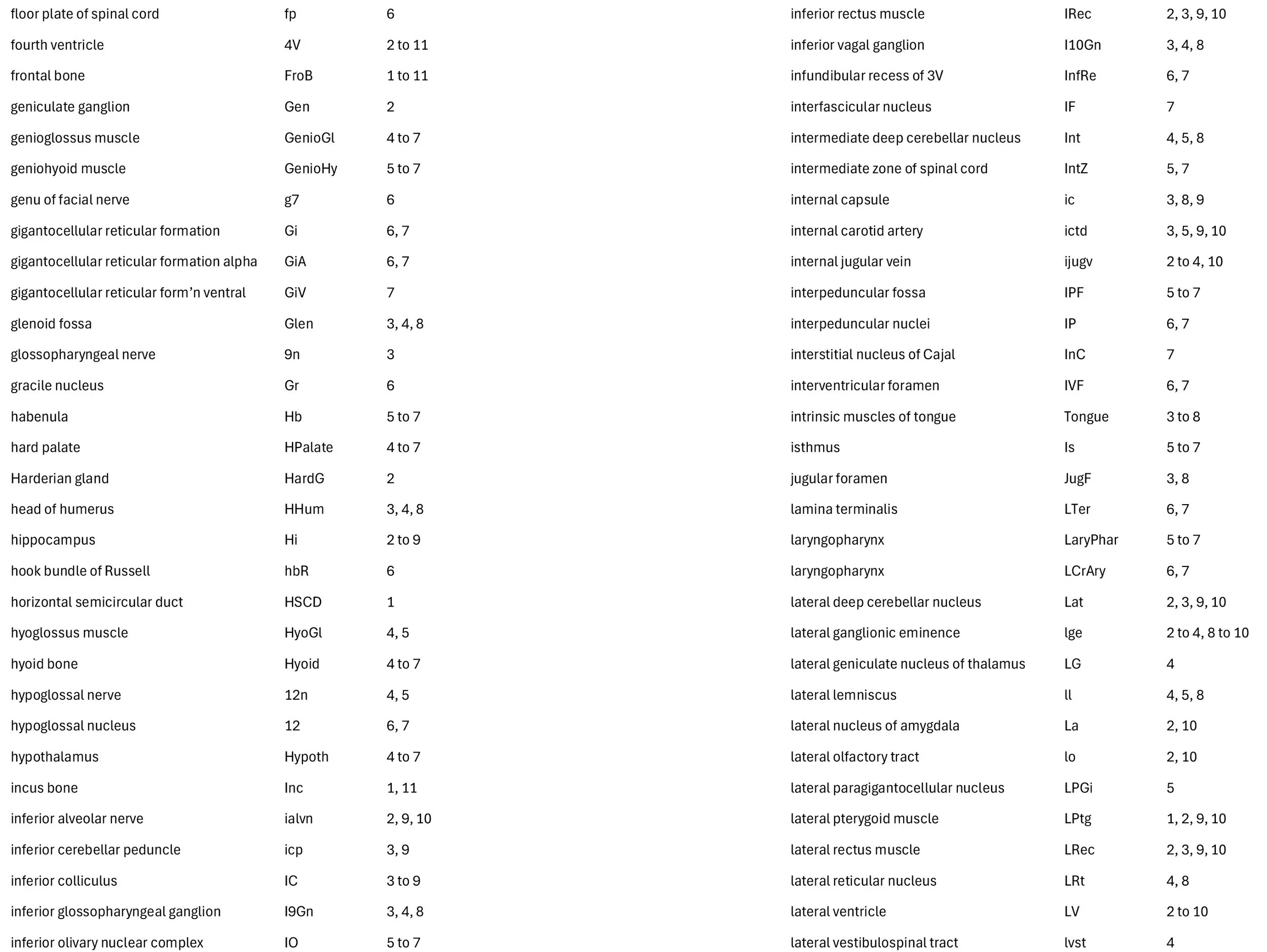
Sagittal Series Through the Head of a Newborn Bandicoot (Perameles nasuta MS216)
Biology of Perameles nasuta
Perameles nasuta (the long-nosed bandicoot) is found along the Eastern Australian coast from Queensland to Victoria. Adult body weight is about 700 g and the nocturnal species is omnivorous, subsisting on soil-dwelling invertebrates, fungi and plants. It’s most distinctive adult feature is it’s long nose, but at the age of the specimen below the snout is blunt and covered by an oral shield.
Methods
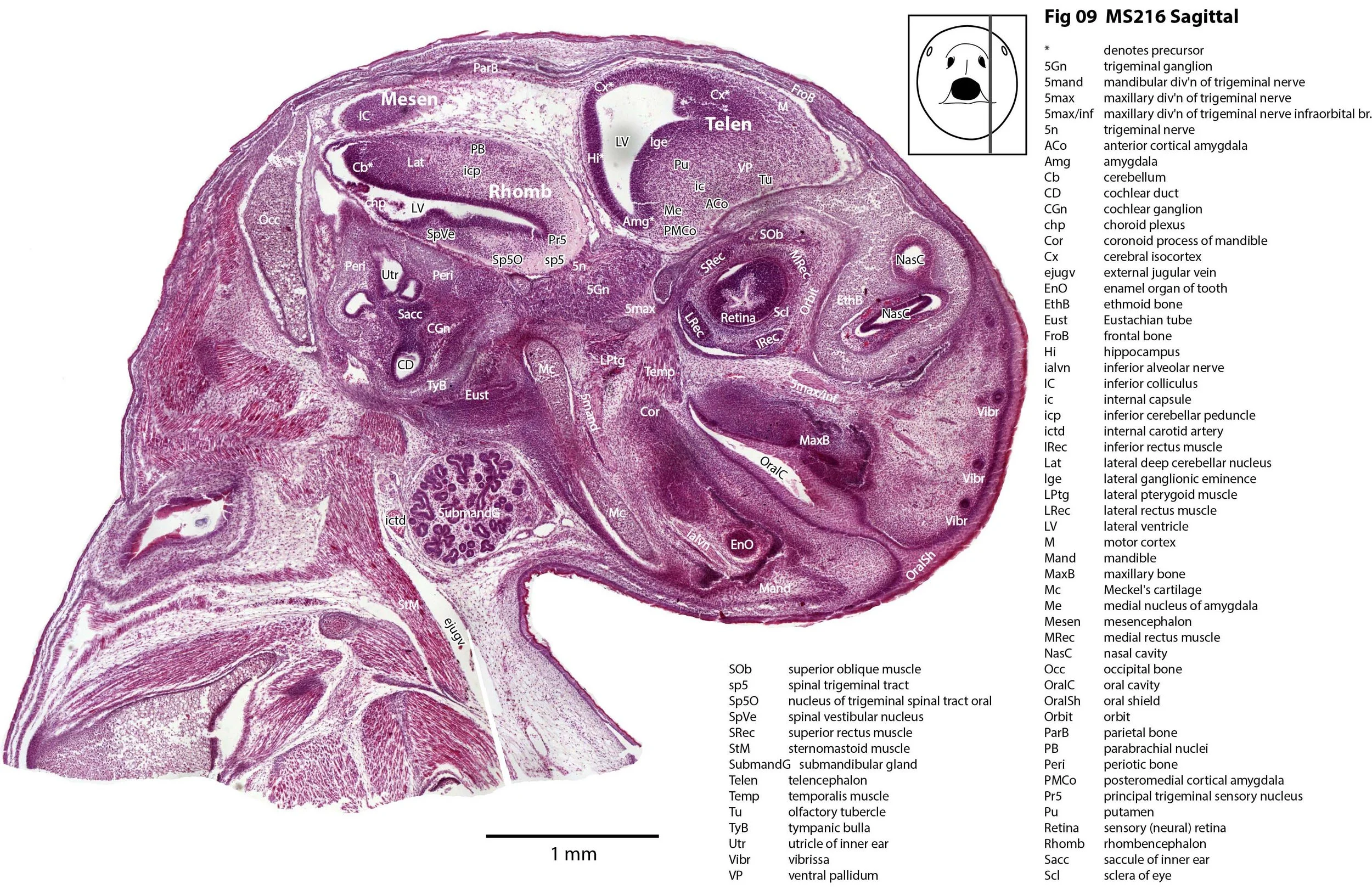
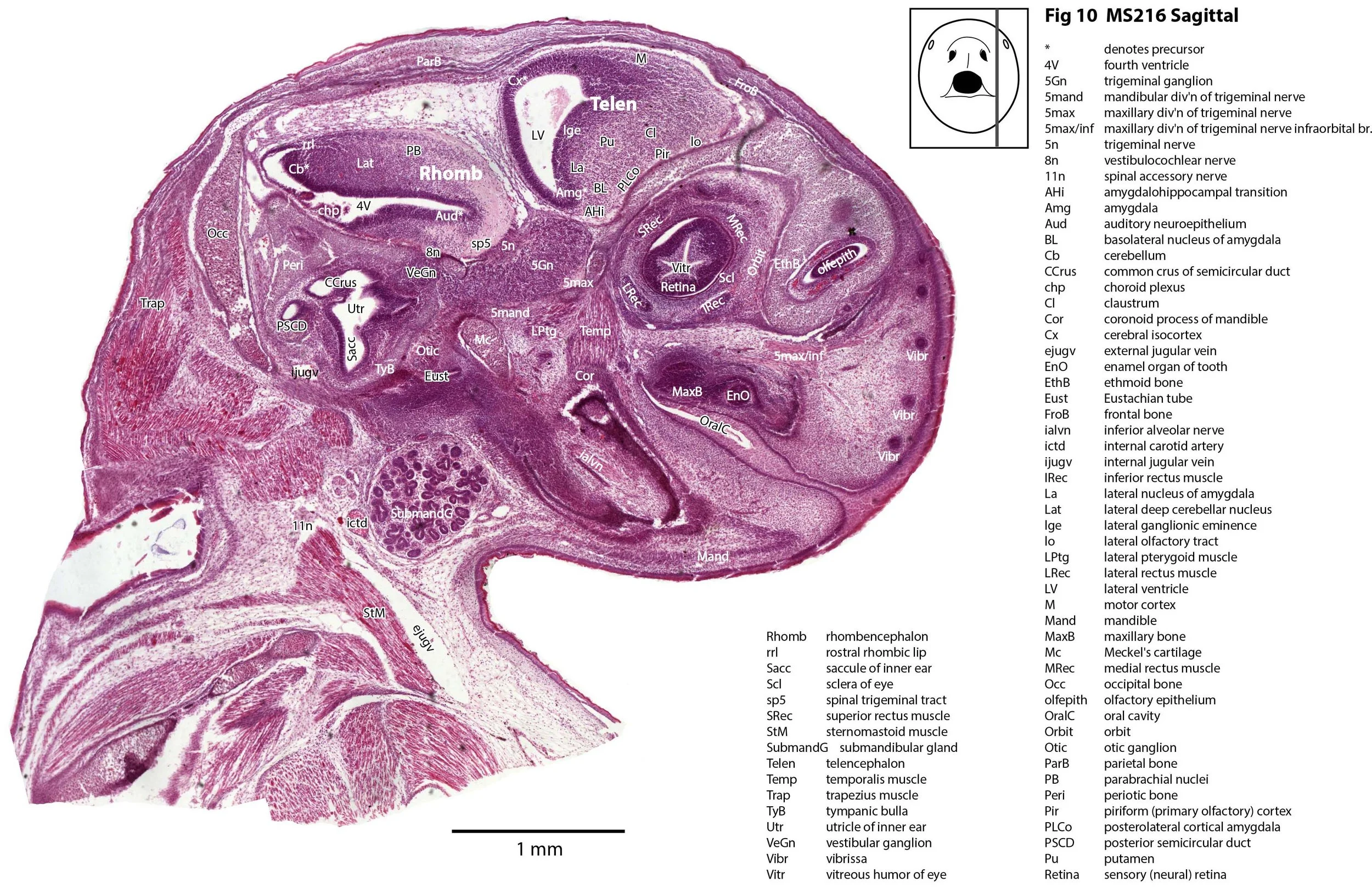
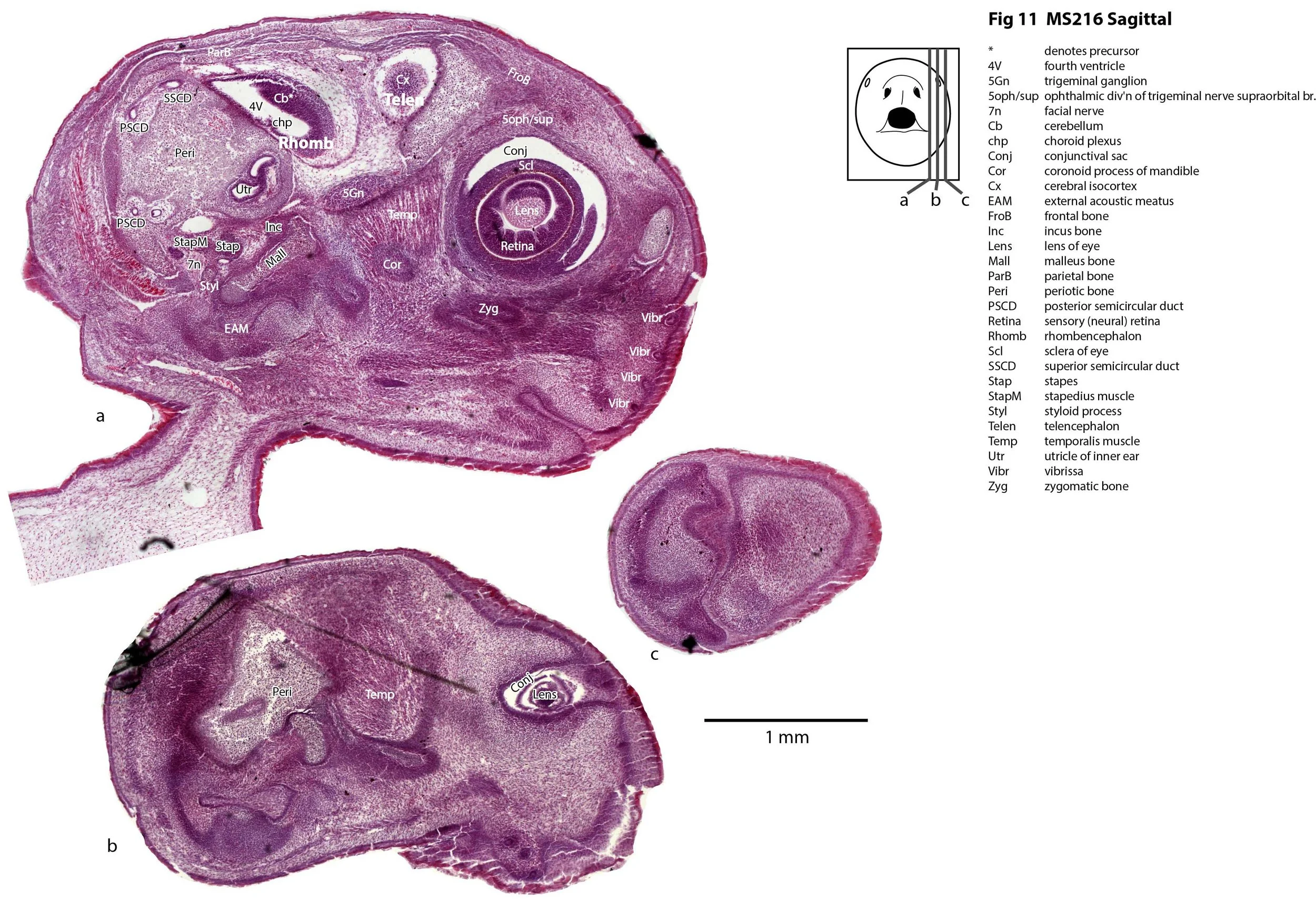
This specimen (listed as Perameles nasuta in Museum records) is part of the JP Hill collection currently at the Museum für Naturkunde Berlin. It had been collected in 1903 and fixed in Zenker’s solution. It is a newborn bandicoot of 14 mm GL and 7 mm HL. The head had been sectioned longitudinally (parasagittally) at a thickness of 10 µm and stained with what appears to be haematoxylin and eosin.
The sections at approximately 220 µm intervals were photographed with the aid of a Zeiss Axioplan2 fitted with an AxioCam MRc5 camera. All images were calibrated by photographing a scale bar at the same magnification. Images were placed in Adobe Illustrator 2021 and delineated. Developmental regions (i.e. neuroepithelium) destined to give rise to adult structures have been denoted by the adult structure’s name with an asterisk (e.g. Cx* denotes the developmental field of the cerebral isocortex).
Observations
The observations that can be made on this specimen are broadly similar to those already made for MS221 (bandicoot newborn frontally sectioned head). In general, the central nervous system, cranial skeleton and craniofacial musculature in newborn bandicoots are intermediate in differentiation between those features in a newborn dasyurid (e.g. quoll) and a newborn macropod (e.g. tammar wallaby).
The nasal cavity has distinct respiratory and olfactory epithelium (plates 2 to 5, 8 to 10), but the olfactory bulb (OB in plates 3 to 7) is still in a primitive state without clear lamination. The piriform (primary olfactory) cortex (Pir in plates 2, 10) is still very immature and poorly laminated.
The rest of the telencephalon consists of an immature cerebral isocortex (Cx in plates 2 to 11), a poorly defined hippocampal inflexure (Hi in plates 2 to 9), and immature basal ganglia (Cd, Pu) receiving neurons from the lateral and medial ganglionic eminences (lge, mge in plates 2 to 5, 8 to 10).
The hypothalamus (plates 4 to 7) is mostly neuroepithelium surrounding the rostral (ventral) third ventricle. Optic (OptRe in plates 6 and 7), infundibular (InfRe in plates 6, 7) and mammillary (MRe in plate 7) recesses of the third ventricle are visible.
The diencephalon (plates 4 to 7) has three distinct prosomeres (p1, 2, 3) arranged caudorostrally. Postmitotic neuronal populations are minimal and it is difficult to discern any thalamic nuclei apart from the LG (plate 4).
The brainstem is more advanced in this species at birth than the newborn dasyurid, but not as differentiated as the newborn tammar wallaby. The trigeminal sensory nuclear complex (Pr5, Sp5O, Sp5I, Sp5C; plates 3 to 9) is easily distinguished. This would be consistent with a major role for trigeminal somatosensation in analysing the environment and feeding information to the brainstem reticulospinal pathways for co-ordination of forelimb movement.
The cerebellum is of a similar level of differentiation to an E14 rat (plates 2 to 11), with very few postmitotic cells and only rudimentary inferior and superior cerebellar peduncles (plates 3, 7, 9). Precerebellar nuclei are also rudimentary, with the inferior olivary complex (IO in plate 5 to 7) present, but not pontine nuclei.
The spinal cord is most differentiated in the cervical region (e.g. plates 5 to 7), but even here there is very limited differentiation of the dorsal (sensory) horn.
Acknowledgements
I would like to thank Dr Peter Giere of the MfN, Berlin Germany, for access to the collection and for all his kind help during the work.
References
Diogo R, Bello-Hellegouarch G, Kohlsdorf T, Esteve-Altava B, Molnar JL (2016) Comparative myology and evolution of marsupials and other vertebrates with notes on complexity, bauplan, and “scala naturae”. Anat Rec 299, 1224-1255.
Schneider NY, Gurovich Y (2017) Morphology and evolution of the oral shield in marsupial neonates including the newborn monito del monte (Dromiciops gliroides, Marsupialia Microbiotheria) pouch young. J Anat 231, 59-83.